A few SGers were confused by the way in which the 2009 paper for the e. coli experiment didn’t provide clear numbers regarding what was, and what was not, a fixation event. And, as it turns out, considerably more work has been done in this space since 2009, and it’s evident that at least a few scientists are growing concerned about the obvious implications of the time, in generations, that is required for mutational fixation, presumably due to its obvious implications for the impossibility of evolution by natural selection. In fact, some are even attempting to prove that fixation is not required for adaptation, in a quixotic illustration of the way in which they still don’t truly understand the extant nature of the problem MITTENS poses for TENS.
Any understanding of an evolutionary process requires the understanding of the particular components that make up that evolutionary process. Darwin wrote about these evolutionary processes in his book, “On the Origin of Species”[1]. From this text, we get a particular quote that describes these processes: “For it should be remembered that the competition will generally be most severe between those forms which are most nearly related to each other in habits, constitution and structure. Hence all the intermediate forms between the earlier and later states, that is between the less and more improved state of a species, as well as the original parent-species itself, will generally tend to become extinct. So it probably will be with many whole collateral lines of descent, which will be conquered by later and improved lines of descent. If, however, the modified offspring of a species get into some distinct country, or become quickly adapted to some quite new station, in which child and parent do not come into competition, both may continue to exist.” Darwin recognized that two processes can occur during evolution, competition (what Darwin also calls the struggle for existence) and adaptation. Many papers have been written about the mathematics of competition. Some of the many examples were written by Haldane et al. [2-5]. Here, we will address both the mathematics of competition and the mathematics of adaptation. In this paper, we consider a particular experimental evolutionary model, the Lenski E. coli long term evolution experiment (LTEE) [6] and the particular evolutionary components which cause the experiment to act in its manner. And to address why it takes so many generations for each fixation and adaptation step. A model for fixation was presented in the following paper BH Good, et al. and edited by Richard Lenski where they discuss these issues [7]…
In this study, we make the simplifying assumption that the distribution of variants in the entire 10ml test tube is identical to the distribution of variants in the 0.1ml sample. The other selection process is the natural selection process that occurs as the different variants replicate where the more fit variant ultimately substitutes for the less fit variants over generations. This is due to the differences in the relative fitness of the different variants in the population. These two selection processes must be combined to correctly describe the fixation process in the LTEE. But the fixation process does not describe the adaptation process. Adaptation (an improvement in fitness) occurs when another beneficial mutation occurs on the more fit variant. The improvement in fitness of a particular variant is dependent on the absolute fitness of that variant to replicate because the random trial for improvement in fitness is the replication and the frequency at which the beneficial mutation occurs is given by the mutation rate. This is a binomial probability problem, that is, does the beneficial mutation occur or does it not occur with that replication. The mathematical model to describe this part of the evolutionary process is given by the mathematics of random mutation and natural selection [10]. The probability of that beneficial mutation occurring on the more fit variant is dependent on the number of replications of that variant. This value will be tabulated in the mathematical model of fixation. The key point to understand in the evolutionary process is that fixation is not a requirement for adaptation. If the carrying capacity of the environment is sufficient to allow for the number of replications required to give a reasonable probability of a beneficial mutation occurring on some variant, then fixation is not needed for adaptation.
Have you spotted the problem yet? Anyhow, the experiment provided a way to test how long it took a mutation to fixate, and the results were considerably faster than the 1,600 generations previously indicated. In fact, they managed to reduce the number of generations to fixation to a range between 119 and 497!
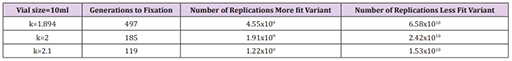
However, the table raises an obvious question. What is this k that appears to be directly related to the number of generations required? And aye, there’s the rub.
If that weight factor is 2, that means that variant is doubling in number every generation.
So, k=2 is a very strong replication weight factor. And yet, even with it cranked up to an unnaturally high rate, it’s still insufficient to reduce the number of generations to fixation to a sufficient level. And a more realistic replication weight factor that is more reflective of the genetic changes we can observe in real-time from one generation to the next is obviously going to be much lower, and in line with those previously reported.
But all of this frantic activity to prove that fixation is not necessary for adaptation is misplaced anyhow, because the problem is not adaptation, but the observable genetic distance that consists of historical fixation events separating one species from another. One can certainly prove that it’s not necessary to travel from point A to point B in a boat in order to cross a river, but that proof will never change the previously measured width of the river.